Introduction
Sierra Nevada Mountain Yellow Legged Frogs (Rana sierrae) were historically one of the most abundant species in Sierra Nevada lakes and streams. However, populations have been decimated by predation by non-native fish, climate change, pesticide drift, the spread of chytriomycota, and habitat fragmentation (Bonham et al. 2011). R sierrae is now federally listed as an endangered species. Desire to support the recovery of this species has motivated examination of the potential for reintroduction of frogs to their historic range. In this case study, we assess Tuolumne Meadows in Yosemite National Park as a potential site for reintroduction. Although R. sierrae is known to have historically occupied Tuolumne Meadows, recent surveys have failed to identify any populations in the area (Brown et al. 2014). Despite this, we hypothesize that Tuolumne Meadows would be a good location to reintroduce the R. sierrae species because of the lentic habitat it could provide. To assess this hypothesis, analysis of habitat suitability was determined through analysis of discharge, water year type, historical imagery, fish surveys, and macroinvertebrate surveys.
Background
Ecology
The historic range of R. sierrae encompassed the entirety of the Sierra Nevada mountain range between the Feather River and the Monarch Divide and Cirque Crest in elevations of approximately 1400 m to 3690 m (Brown et al. 2014). R. sierrae prefer lentic habitat, occupying lakes, ponds, marshes, and low gradient streams. The species overwinters in non-frozen water refuges found in lakes, stream beds, banks, springs, and nearshore bedrock crevices at all lifestages. In the summer adults utilize open shorelines to bask and forage for aquatic macroinvertebrates, terrestrial invertebrates, and tadpoles of other amphibians; while tadpoles consume algae and diatoms. Adult frogs may occupy ephemeral habitat in the early summer to forage and then move to permanent habitat in the fall (Bonham et al. 2011).
Breeding begins at snowmelt and takes place in lakes and ponds with depths greater than four meters or in low gradient stretches of perennial streams. Because of the brief growing season in higher elevations, R. Sierrae tadpoles require 2 to 3 summers before metamorphosis. Tadpole are vulnerable to mortality by anoxic conditions created by complete freezing of a water body or habitat desiccation. Historically, R. sierrae had few aquatic predators and existed in a fishless habitat (Bonham et al. 2011).
Main Threats
Once described as one of the most common species in the Sierra Nevada (Gindell and Storier 1924), remaining R. sierrae populations are alarmingly small, often consisting of isolated populations containing fewer than 20 individuals (Matthews and Miaud 2007). It is hypothesized that fish introductions have been the main driver of R. sierrae population declines (Knapp, 1996). Fish also overwinter in non-frozen refuges, and heavily prey upon R. sierrae to sustain themselves through the winter. In addition, R. sierrae populations have been negatively impacted by climate change, pesticide drift, the spread of chytridiomycota, and the resulting habitat loss and fragmentation (Brown et al. 2014).
Tuolumne Meadows
Tuolumne Meadows is one of the largest sub-alpine meadows in the Sierra Nevada. It is at an elevation of approximately 2600 m and is characterized by a thin layer of topsoil underlain by granitic bedrock. Here, meadow hydrology is dominated by snowmelt and precipitation. Despite records of the historic presence of R. sierrae in Tuolumne Meadows, surveys conducted from 1995-2001 noted the absence of any frog species (Brown et al. 2014). Nearby Kuna Lake and the Conness Lakes support current populations of R. sierrae, although these sub-populations are isolated in larger, high alpine, lentic environments with little to no population connectivity with the surrounding meadows (Brown et al. 2014, Matthews and Miaud 2007).
Methods
The suitability of Tuolumne Meadows was analyzed through the collection of historic imagery, discharge, fish surveys, water year type, and macroinvertebrate surveys. To quantify aquatic habitat availability and persistence, discharge and backwater areas were analyzed. Discharge data from the Tuolumne River in Tuolumne Meadows was retrieved from California Department of Water Resources (DWR) and averaged by month. Mean monthly discharge was plotted with error bars to compare seasonal variability. Area of lentic backwater and side- channel stream habitat in June was quantified for 2009, 2012, 2013, and 2014 using historic imagery of the meadows from Google Earth Pro. Water year type was obtained from the San Joaquin Water Index to classify each year from 1995 to 2015 as wet, above average, below average, dry, or critically dry. This classification was also used to relate the amount of water the meadow received and to the change in the amount of backwater and side channel habitat.
Fish snorkel survey data at low flow was obtained from previous years of the undergraduate ecogeomorphology class, conducted by the Center for Watershed Science at UC Davis (UCD CWS).
Aquatic invertebrate samples were collected to quantify stream health and potential food availability for frogs in June 2016. Due to high flows, surveys for macroinvertebrates were conducted in three small Tuolumne River tributaries; Ecogeo Creek, Budd Creek, and Unicorn Creek. Composite kick samples, comprising of three sixty-second kick intervals, were collected at three randomly selected transects per reach. Captured macroinvertebrates were sorted in the field by order, and stream health was calculated by the Hilsenhoff Biotic Index where a score of 0 indicates excellent water quality with no organic pollution and 10 indicates very poor water quality with severe organic pollution (Hilsenhoff, 1987).
Results
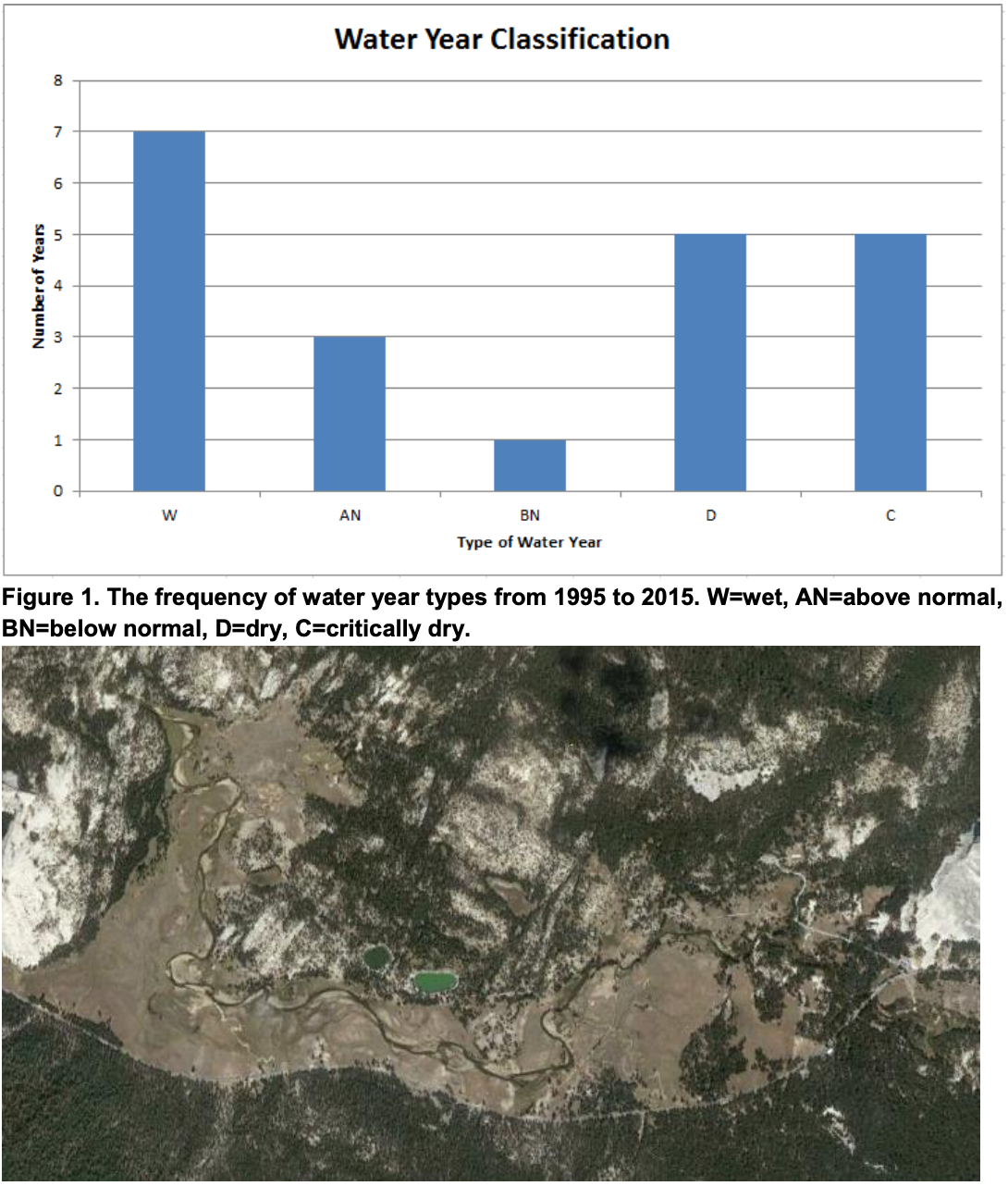
Analysis of the mainstem Tuolumne River showed the stream gradient through Tuolumne meadows to be 4.46 ft/mi. The number of wet, above normal, below normal, dry, and critically dry water years are quantified in Figure 1, and show that almost half of water years are dry or critically dry. Variability in monthly discharge was compared for the last three years of drought (Figure 4), which showed that deviations from monthly mean discharge were most extreme in the winter and early spring months and relatively small in the summer months.
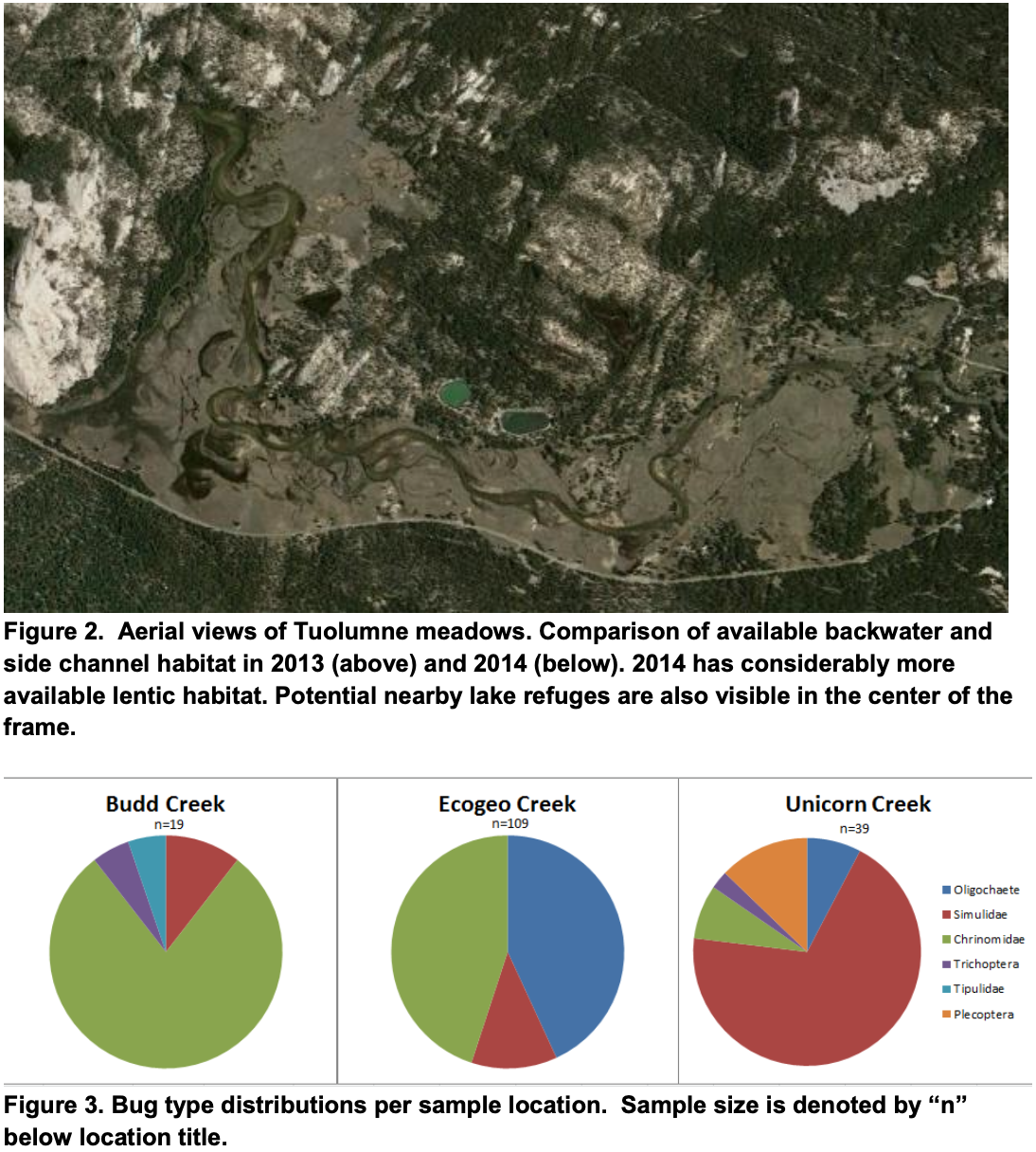
Comparing the summer habitat availability in a dry versus drought year in Google Earth Pro revealed the presence of habitable backwater habitat. Figure 2 shows the contrast of a dry year (2009) to a critically dry year (2013). It can be seen that there is a stark difference in the habitat available due to drought. Table 1 shows the area of habitat available outside of the main channel in each water year type. This analysis showed that replenishment of backwater habitat is not necessarily correlated to water year type, and lentic habitat is available even in prolonged periods of drought.
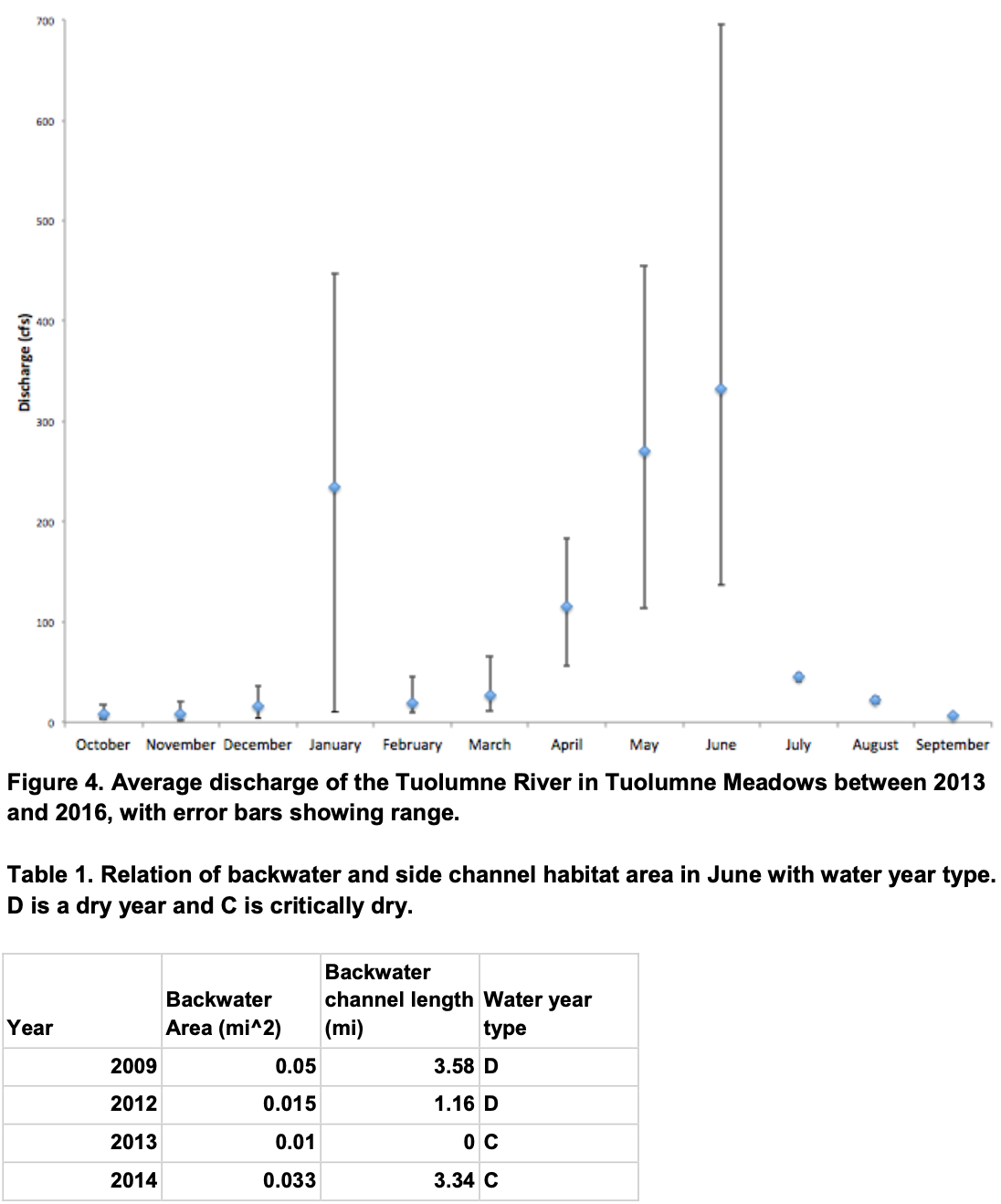
Benthic macroinvertebrates were collected in Budd, Unicorn, and Ecogeo Creek (Figure 3) and were used to calculate HBIs through the use of Hilsenhoff’s assigned values of pollution tolerance for each macroinvertebrate order. This calculation yields a ratio of tolerant species per reach and gives an estimate of water quality and pollutant contamination. Budd and Unicorn Creek were both found to have HBI values of approximately 5, indicating good water quality and some organic pollution. EcoGeo Creek had a slightly higher HBI value of 6, indicating slightly lower water quality and an increase in organic pollution.
Discussion
Analysis of the hydrology of Tuolumne Meadows showed high seasonal and annual variability. Inflow of water to Tuolumne Meadows is dominated by snowmelt and precipitation due to its high elevation and shallow topsoil, which means that the amount of water in the meadow is highly dependent on climate patterns and water year type. Figure 1 shows that dry or critically dry years are common to the watershed, and should be expected almost half of the time. However, our analysis of the main channel showed that discharge during the summer months was relatively predictable despite large variation in winter flows. As a result, the main channel may serve as a refuge for frogs during drought because of its reliable perennial flow and low gradient. Further, there are two perennial lakes just above the meadows that are accessible enough to be additional refuges in critically dry years (Figure 2).
Analysis of historical imagery indicates that the availability of backwater habitat is not necessarily dependent on water year type. Despite the ongoing drought in 2013 and 2014 (both years are critically dry), there was a large increase in area of backwater habitat between these years (Table 1) indicating that flooding of the meadow is more dependent on individual precipitation or snowmelt events than total annual discharge. Therefore, even if droughts increase in frequency and severity, lentic habitat could be maintained for frogs by a few large flow events over the winter.
Current climate change models predict an overall decrease in snowpack throughout the Sierra Nevada and an increase in rain events (Cayan et al. 2008). The snowpack that supplies the Tuolumne is expected to be more persistent than average due to the high altitude of the Southern Sierras. This suggests that the Tuolumne watershed will be less affected by climate change than other Sierra Nevada watersheds, and thus more likely to support R. sierrae populations in the future compared to other stream systems. If the watershed becomes more dominated by rain events than snow, peak discharge is expected to increase (Cayan et al. 2008). Increased stream flooding could replenish lentic habitat in the meadow via overbank flows and support the persistence of R. sierrae populations. Therefore, Tuolumne Meadows could provide the necessary habitat for long term persistence of R. sierrae populations.
Macroinvertebrate surveys show evidence of clean aquatic habitat and sufficient food availability for R. sierrae in Tuolumne Meadows (Figure 3). HBI index values indicate good water quality with limited pollution. Low levels of pollution are likely due to the meadow’s close proximity to State Route 120, portable toilets, campgrounds, a store, and many trails and overlooks. However, the park is actively seeking adaptive management and restoration strategies to improve meadow conditions. Therefore, the HBI index may decrease with time.
Although these metrics indicate that Tuolumne Meadows could support a reintroduced R. sierrae population, immediate reestablishment of populations would be unsuccessful due to the presence of fish and a lack of connectivity to other existing amphibian populations. Fish snorkel surveys in previous years by UCD CWS found populations of Rainbow Trout (Oncorhynchus mykiss), Brown Trout (Salmo trutta), and Brook Trout (Salvelinus fontinalis) in Tuolumne Meadows. High rates of predation and competition for resources of these fishes with R. Sierrae would make it impossible to reintroduce a self sustaining population of frogs (Vrendenburg et al. 2004). Tuolumne meadows is also connected to fish-containing streams and lakes via the Tuolumne River and its tributaries. Therefore an extensive removal program would have to take place and a physical or physiological barrier would have to be established on the eastern boundary of the meadows in order to keep the habitat permanently fishless.
If the fish removal programs were successful, isolation from surrounding populations might make the reintroduced population of R. sierrae vulnerable to a genetic bottleneck. Despite presence of nearby populations in surrounding higher alpine lakes, reintroduced populations may be physically isolated from existing populations due to large differences in elevation. This isolation may result in low genetic variability due to a lack of genetic drift between populations. As a result, the population genetics may become homogenous, providing low genetic resistance and decreasing the population’s resilience to future threats. Therefore, despite adequate habitat suitability, R. sierrae should not be reintroduced to Tuolumne Meadows until fish are removed and connectivity is established.
Conclusion
The Sierra Nevada Yellow Legged frog is critically endangered and absent from an overwhelming majority of its historical range. This drastic collapse of a species once abundant throughout the northern Sierra Nevada necessitates action to prevent extinction. Reintroduction of R. sierrae to historically occupied areas could be an immediate and effective way to support the recovery of this species. We examined Tuolumne Meadows in Yosemite National Park as a possible site for reintroduction of R sierrae by analysis of discharge, water year type, historical imagery, fish surveys, and macroinvertebrate surveys. Our analysis showed that Tuolumne meadows could indeed support a viable population of R sierrae due to lentic habitat availability, good stream health, and food availability. However, currently extensive invasive fish would decimate reintroduced populations by predation and competition, and a lack of connectivity to other frog populations could result in genetic bottlenecking. Restoration efforts for R. sierrae should therefore take a headwater-down approach, where fish are systematically removed and frog populations restored from the top of the watershed down to the meadows. This would reestablish population connectivity, and provide R. sierrae with high potential for recovery.
Works Cited
Bonham, C. 2011. A status review of the Mountain yellow-legged frog. California Department of Fish and Game, State of California Natural Resources Agency.
Brown, C. H., G. Green, and D. Macfarlane. 2014. Mountain yellow legged frog conservation assessment for Sierra Nevada, CA. Pacific Southwest Regional Office, USDA Forest Service U.S. Fish and Wildlife Service: 138
Cayan, D. R., E. P. Maurer, M. D. Dettinger, M. Tyree, and K. Hayhoe. 2008. Climate change scenarios for the California region. Climatic Change 87.S1 (2008): 21-42. Web
Grinnell, J., and T. I. Storer. 1924. Animal life in the Yosemite. University Press.
Hilsenhoff, W. L. 1987. An improved biotic index of organic stream pollution. Great Lakes Entomologist 20.1: 31-40.
Knapp, R. A. 1996. Non-native trout in natural lakes of the Sierra Nevada: An analysis of their distribution and impacts on native aquatic biota. Sierra Nevada Ecosystem Project. Davis: University of California, Centers for Water and Wildland Resources, University of California. III.
Mattews, K. R., and C. Miaud. 2007. A skeletochronological study of the age structure, growth, and longevity of the Mountain yellow-legged frog, Rena muscosa, in the Sierra Nevada, California. Copeia:986-993
Vredenburg, V. T. 2004. Reversing introduced species effects: Experimental removal of introduced trout leads to rapid recovery of a declining frog. Ecological Society of America Annual Meeting Abstracts 89: 526.